- OSMORÉGULATION
- OSMORÉGULATIONIl est couramment admis que la vie sur la Terre a pris naissance dans le milieu marin et que les autres biotopes (eaux douces, saumâtres, sursalées; sol; atmosphère terrestre) ont été colonisés ultérieurement par les êtres vivants. Ces divers environnements se distinguent les uns des autres principalement par leur teneur en eau et en sels. Or ces dernières substances sont aussi des constituants fondamentaux de la matière vivante; elles sont responsables pour une très large part de la pression osmotique des liquides internes, qui est une caractéristique biologique importante.L’ensemble des mécanismes par lesquels les organismes effectuent le contrôle de leur pression osmotique interne constitue l’osmorégulation, qui revêt des formes très variées et dont l’étude, actuellement bien documentée, constitue un chapitre passionnant de la physiologie comparée.1. Importance de la pression osmotiqueChez les êtres unicellulaires, les échanges avec le milieu externe s’effectuent directement à travers la membrane cellulaire. Chez les organismes pluricellulaires évolués, Claude Bernard avait montré qu’un «milieu intérieur» constitué par les fluides internes (sang, lymphe, liquides interstitiels) et interposé entre les cellules et le milieu ambiant assurait par la «fixité» de sa composition des conditions physiologiques favorables [cf. HOMÉOSTASIE]. Cette dernière conception a dû être grandement révisée; il est plus exact de dire que la composition des fluides extracellulaires est relativement stable, ou n’est sujette qu’à des variations limitées et compatibles avec la vie cellulaire.Les liquides externes et internes sont constitués par de l’eau tenant en dissolution des substances minérales (essentiellement les ions sodium Na + et chlore Cl-) et organiques. Ces substances, se comportant dans l’eau comme des molécules de gaz dans une atmosphère gazeuse, exercent sur les parois des cellules une pression, la pression osmotique P, qui peut être exprimée en atmosphères (pour 1 litre, à 0 0C). À présent, il est courant de l’indiquer en osmoles par litre ou par kilogramme d’eau, par comparaison avec des solutions étalons de chlorure de sodium. Ainsi, une solution contenant une mole de glucose, ou 180 g/l, possède une concentration de 1 osmole par litre: une solution contenant 1 mole (58,5 g) de chlorure de sodium par litre libère deux ions par molécule et possède une concentration osmolaire de 2 osmoles par litre (en pratique, il faut tenir compte d’un facteur de dissociation, donné par des tables). La concentration totale de ces fluides peut encore être mesurée d’après l’abaissement du point de congélation ( ) ou abaissement cryoscopique , en degrés Celsius. Approximativement:
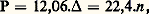 où n est le nombre d’osmoles par litre. Le tableau donne quelques valeurs caractéristiques de la concentration osmotique.Une différence de pression osmotique de part et d’autre d’une membrane perméable entraîne au moins un déplacement d’eau (transport osmotique), du milieu le moins concentré vers le plus concentré; éventuellement, les sels dissous migrent en sens inverse jusqu’à ce que l’équilibre soit établi entre les deux côtés. Au vu du tableau, il est évident que de tels mouvements doivent exister chez les êtres vivants.2. Perméabilité des membranes biologiquesLes échanges osmorégulateurs sont assurés chez les organismes vivants par des organes perméables spécialisés: tégument, branchies, tube digestif, poumons, reins, etc., constitués par des épithéliums simples ou pluristratifiés, que l’on qualifie indistinctement de membranes biologiques (ce terme englobant en fait, également, les membranes cellulaires). On ne peut entrer ici dans le détail de l’importante question de leur perméabilité, dont les propriétés fondamentales ont été mises en évidence par des travaux effectués sur la cellule végétale (W. Pfeffer, C. E. Overton). Les parois ne sont pas semi-perméables, c’est-à-dire qu’elles se laissent traverser à la fois par l’eau et par les substances dissoutes; mais leur perméabilité est très sélective; ainsi, les ions Na+ et Cl- sont essentiellement extracellulaires, tandis que le potassium (K+) est surtout intracellulaire. L’utilisation, à présent généralisée, des marqueurs radioactifs met en évidence que les échanges transmembranaires sont bidirectionnels, c’est-à-dire qu’ils s’effectuent dans les deux sens. Ils sont, en outre, très intenses, ce qui montre que l’eau et les sels des êtres vivants se renouvellent à grande vitesse.Dans certains cas, ces échanges sont passifs , c’est-à-dire que, pour la substance considérée, leur intensité est prévisible d’après des lois physico-chimiques simples: elle est calculable d’après la différence de concentration (gradient chimique) et la différence de potentiel électrique (gradient électrique) existant entre les deux faces; l’équilibre est régipar la loi de Gibbs-Donnan. Dans d’autres cas, l’équilibre que l’on constate de fait apparaît comme un «défi» permanent à ces mêmes lois; il n’est donc maintenu qu’au prix d’une dépense énergétique que la cellule transporteuse effectue en permanence; on dit que le transport est actif . Ce dernier cas correspond à des « pompes biologiques», dont la plus répandue et la plus étudiée est la «pompe à sodium», qui refoule les ions Na+ hors des cellules [cf. MEMBRANES CELLULAIRES].Les mécanismes biochimiques qui régissent ces transports sont encore mal connus. Dans un certain nombre de cas, on sait qu’ils sont contrôlés par des hormones et par des enzymes (telles que les ATPases, qui libèrent de l’énergie aux dépens de l’ATP), assurés par des transporteurs spécifiques, enfin conditionnés par la structure des membranes (replis de la paroi cellulaire, richesse en mitochondries, etc.).3. Types d’osmorégulationL’équilibre est isosmotique lorsqu’il y a égalité de pression osmotique entre le milieu inférieur et le milieu externe. Pratiquement, ce cas ne s’observe que pour des animaux marins. L’organisme ne tend ni à s’hydrater, ni à se déshydrater. La régulation osmotique porte essentiellement sur les mouvements d’ions (ionorégulation), tandis que le transport osmotique de l’eau est pratiquement nul.Dans les autres cas, l’équilibre est anisosmotique . Quand le milieu intérieur est le moins concentré (hypo-osmotique ), l’animal tend à se déshydrater et à être envahi par le sel qui diffuse à travers les surfaces externes. Il doit compenser ces mouvements en absorbant du liquide externe et en excrétant activement l’excès de sel. Au contraire, si le milieu est le plus concentré (hyperosmotique ), l’animal tend à s’hydrater et à perdre du sel passivement; il compense en excrétant l’excédent d’eau et en absorbant activement des ions à partir du milieu environnant.Les animaux sténohalins sont adaptés à un seul milieu et ne supportent en général que de faibles variations de la salinité externe. En revanche, les euryhalins résistent à de larges variations; les plus caractéristiques sont les migrateurs, tels que l’anguille, le saumon et certains crabes, qui peuvent passer alternativement de l’eau douce à l’eau de mer et chez lesquels les mécanismes osmorégulateurs doivent donc s’inverser quand l’animal change de milieu.L’habitat aérien se caractérise par sa teneur en vapeur d’eau atmosphérique (humidité) et non plus par la salinité. Le problème majeur pour l’animal est de réduire, et d’équilibrer convenablement, les pertes d’eau résultant de l’excrétion, de l’évaporation cutanée et de la vaporisation au niveau des organes respiratoires.Ces divers cas sont illustrés ci-dessous au moyen d’exemples pris dans les différents milieux naturels.Osmorégulation en milieu salinPar leur masse, les océans constituent le milieu d’habitat le plus vaste. La composition et les caractères physico-chimiques y sont remarquablement constants [cf. OCÉANS ET MERS]. La salinité est voisine de 34 g de chlorure de sodium pour 1000, ce qui correspond à une osmolarité de l’ordre de 1 osmole par litre. Assez curieusement, bien que les teneurs absolues en ions minéraux soient très variées dans les liquides extracellulaires des animaux, les proportions de ces ions y sont cependant comparables à celles que l’on trouve dans l’eau de mer. Cette constatation est un argument d’ordre physiologique avancé à la fin du siècle dernier en faveur de l’hypothèse de l’origine marine des êtres vivants (F. Botazzi).Les autres milieux salins naturels sont constitués, d’une part, par les eaux saumâtres, dont la salinité, inférieure à celle de la mer, est très variable; d’autre part, par les salines des déserts où, du fait d’une évaporation intense, la concentration est si élevée que le sel précipite.Osmorégulation des InvertébrésLa grande majorité des Invertébrés sont en équilibre isosmotique avec l’eau de mer, mais la composition de leur milieu intérieur en diffère sensiblement et de façon variable suivant les espèces. Ainsi, chez la ligie (Crustacé isopode), le sodium de l’hémolymphe est de 20 p. 100 plus élevé que dans la mer, tandis que le calcium est multiplié par 3 et le magnésium diminué de 60 p. 100. Le tégument et les autres membranes externes sont très perméables à l’eau et aux sels, comme le montrent les études isotopiques; ce fait semble expliquer qu’ils soient isosmotiques. Les capacités osmorégulatrices peuvent être testées en soumettant ces animaux à des changements plus ou moins amples de la salinité. La figure 1 illustre les types fondamentaux d’osmorégulation observés chez le groupe le mieux étudié, celui des Crustacés. Chez les espèces osmoconformes , l’osmolarité interne suit passivement et égale celle du milieu externe (Maia ); ces animaux se laissent traverser par l’eau et le sel; leur tolérance aux changements de salinité n’est limitée que par la résistance cellulaire; il survient en milieu dilué, mais pas en eau douce; ils se comportent dans une certaine mesure comme des osmomètres. Carcinus est osmoconforme en milieu concentré et osmorégulateur en milieu dilué, ce qui lui permet, à la limite, de survivre en eau douce. Pachygrapsus possède une bonne osmorégulation à toutes les salinités. Le cas le plus impressionnant est celui de l’artémie des salines (Artemia salina ) qui maintient une concentration interne basse, même dans des eaux saturées en chlorure de sodium.Osmorégulation des VertébrésLes Cyclostomes et les Élasmobranches sont en équilibre isosmotique avec l’eau de mer, mais suivant des mécanismes différents. Chez les premiers, qui constituent le groupe le plus primitif, la forte concentration interne est réalisée comme chez les Invertébrés, par l’abondance des ions minéraux, surtout Na+ et Cl-. Chez les seconds, ces ions ne comptent que pour un tiers environ de l’osmolarité plasmatique; les deux autres tiers sont assurés, d’une façon très originale, par de l’urée, qui est un déchet normal et non toxique du métabolisme azoté chez les Élasmobranches; l’urée ne traverse pas l’épithélium branchial et n’est pas excrétée par les reins; de ce fait, elle s’accumule dans le milieu intérieur.Les Poissons téléostéens et les Tétrapodes sont tous hypo-osmotiques à l’eau de mer; leur concentration interne, voisine de 300 milliosmoles par litre, n’est que le tiers de celle du milieu externe. Le cas classique du Téléostéen marin, représenté dans la figure 2, constitue un bon exemple de régulation hypo-osmotique en milieu salin. De tels animaux boivent plus ou moins d’eau de mer, soit pour compenser la déshydratation périphérique, soit accidentellement pour prendre leur nourriture. L’eau de mer est en partie absorbée à travers l’épithélium digestif, et avec elle des ions monovalents (Na+, Cl-, K+); les ions divalents (Ca2+, Mg2+, S42-) sont peu absorbés et sont évacués par l’anus. Ce faisant, un excès de sel est introduit dans l’organisme. Seuls les Mammifères marins ont, comme les autres Mammifères, un rein hautement évolué, pourvu d’anses de Henlé, qui leur permet de fabriquer une urine plus concentrée que le plasma, donc d’éliminer ce sel superflu. Les autres groupes de Vertébrés sont pourvus d’un rein plus primitif, sans anses de Henlé, produisant une urine au mieux isosmotique au plasma. Le relais est alors pris par d’autres organes osmorégulateurs, qui excrètent activement les ions indésirables. Chez les Invertébrés et chez les poissons marins, ces organes sont les branchies. Chez les Tétrapodes autres que les Mammifères, cette fonction est assurée par des glandes nasales ou orbitaires logées dans la tête. La sécrétion de ces glandes, plus concentrée que l’eau de mer, s’écoule soit au niveau des yeux, sous forme de «larmes», soit à l’extrémité du bec. Cela explique notamment que les oiseaux marins soient capables de rester longtemps au large, sans accès à l’eau douce (K. Schmidt-Nielsen).Osmorégulation en eau douceLes eaux douces ont une osmolarité négligeable (quelques milliosmoles) et réalisent un milieu d’adaptation éminemment variable dans sa concentration, sa température, etc. Tous les animaux d’eau douce sont hyperosmotiques au milieu et tendent à gagner du poids et perdre des électrolytes. L’ingestion d’eau par le tube digestif est donc nuisible et, de fait, normalement limitée. En revanche, ils évacuent de l’eau sous forme d’une urine généralement abondante et très diluée par rapport au milieu intérieur. En outre, ils absorbent activement des ions à partir du milieu externe par l’intermédiaire d’organes perméables spécialisés: branchies, papilles anales de larves d’insectes... Un cas typique de cette osmorégulation est celui du téléostéen d’eau douce (fig. 2).Le cas des poissons migrateurs a suscité depuis longtemps beaucoup d’intérêt, notamment parce que leur épithélium branchial peut effectuer suivant le milieu soit une excrétion, soit une absorption des ions, toutes les deux actives. Le déterminisme de cette inversion du transport n’est, à vrai dire, guère élucidé. Ces échanges semblent se produire au niveau de cellules particulières (dont les «cellules à chlorures»), paraissent dépendre d’enzymes membranaires et sont contrôlés par des hormones de plusieurs glandes (hypophyse antérieure, neurohypophyse, hormones stéroïdes, thyroïde).Osmorégulation en milieu aérienEn milieu aérien, la difficulté principale à surmonter réside dans la déperdition d’eau vers l’atmosphère, due à l’évaporation tégumentaire et aux diverses formes d’excrétion. Ces pertes d’eau atteignent leur maximum dans les déserts, où l’air est particulièrement sec. Les surfaces respiratoires, qui sont relativement vastes et, de ce fait, sujettes à l’évaporation, se recouvrent de replis tégumentaires ou s’enfoncent à l’intérieur du corps (Insectes, Vertébrés). Les pertes sont compensées par l’eau de boisson, dont l’ingestion obéit à un déterminisme physiologique rigoureux (réflexe de la soif). Mais l’eau n’est pas toujours disponible à volonté, d’où la nécessité de palliatifs. Certains insectes (Thermobia ) possèdent la propriété remarquable d’absorber activement de l’eau-vapeur atmosphérique, même dans un milieu relativement sec (50 p. 100 d’humidité relative). Cela suppose un travail «contre-osmotique» considérable, dont le mécanisme fait encore problème. D’autres font appel à l’eau métabolique qui se forme par oxydation intracellulaire des substrats énergétiques (par exemple à partir du glucose):C6 H12 6 + 626C2 + 6H2O(cf. MÉTABOLISME, OXYDORÉDUCTIONS BIOLOGIQUES). Chez les Vertébrés, les Amphibiens restent inféodés à l’eau; leur peau nue étant perméable, ils accumulent une quantité importante d’urine très diluée dans leur vessie lorsqu’ils vivent en milieu sec; cette eau de réserve peut être réabsorbée en cas de sécheresse.Les Reptiles et les Oiseaux sont pourvus de phanères épais et diminuent leurs pertes d’eau en excrétant une faible quantité d’urine; cette dernière particularité tient au fait que, chez ces animaux, le métabolisme des matières azotées n’aboutit pas à la formation d’urée (dont l’excrétion entraînerait beaucoup d’eau), mais d’acide urique, qui précipite et donne à l’urine son aspect blanchâtre et sa consistance presque solide.Les Mammifères, enfin, dont le rein est pourvu d’anses de Henlé, peuvent seuls produire une urine au moins deux fois plus concentrée que le plasma; cela leur permet, d’une part, de se débarrasser de leur sel en excès, d’autre part, d’excréter de l’urée comme déchet azoté, donc d’avoir une urine relativement abondante sans inconvénient pour leur équilibre osmotique. Le cas le plus spectaculaire est celui des Mammifères désertiques (le rat-kangourou et le chameau).Le petit rongeur américain Dipodomys , ou rat-kangourou, vit sans eau alimentaire. L’apport d’eau est constitué par l’eau métabolique produite par la combustion des réserves organiques du corps. L’utilisation de 1 g de graisse libère 1,1 ml d’eau. Les pertes périphériques d’eau sont très limitées, d’une part parce que cet animal ne possède pas de glandes sudoripares, d’autre part parce qu’il évite la chaleur du jour et ne cherche sa nourriture que par les nuits fraîches. Son rein pourvu d’anses de Henlé très longues fabrique une urine extrêmement concentrée puisqu’elle atteint 6 osmoles par litre, soit environ = 漣 11 0C. Un tel animal peut boire de l’eau de mer sans aucun inconvénient, ce que l’homme ne peut faire.Le chameau demeure l’exemple le plus populaire. Il peut rester des jours sans boire; à l’inverse, il est capable d’ingurgiter en une seule fois l’équivalent du tiers de son poids en eau. Contrairement à ce qu’affirme la légende, il n’emmagasine de l’eau ni dans sa bosse, ni dans son rumen, ni dans d’autres organes. La bosse est formée de graisses qui, oxydées, libèrent de l’eau métabolique. Mais cette eau n’est pas de grande utilité, car le chameau a une activité diurne. En fait, les performances exceptionnelles du chameau proviennent du fait que cet animal peut tolérer d’une part une forte déshydratation, d’autre part une élévation de plusieurs degrés de sa température interne. Son osmorégulation et sa thermorégulation sont relativement labiles, ce qui explique sa résistance. Ce dernier exemple, emprunté à un organisme pourtant très évolué, montre bien que l’osmorégulation peut fort bien s’accommoder d’assez larges variations des concentrations internes sans que la survie soit compromise.osmorégulation [ɔsmoʀegylɑsjɔ̃] n. f.ÉTYM. Av. 1969; attesté en angl. en 1957; de 2. osmo-, et régulation.❖♦ Physiol. Ensemble des mécanismes par lesquels les organismes maintiennent leur pression osmotique interne dans un équilibre déterminé (qui peut être isosmotique ou anisosmotique) par rapport à celle du milieu externe. || Osmorégulation et ionorégulation. || Osmorégulation intracellulaire. → 2. Osmorégulateur (cit.).0 Chez les Reptiles et les Oiseaux marins, dont le rein est incapable de produire une urine fortement hypertonique et qui cependant ne disposent comme source d'eau et pendant de longues périodes que d'eau de mer, très fortement hypertonique au plasma sanguin, existe une glande originale, apparemment étroitement spécialisée dans les fonctions d'osmorégulation, la glande nasale (K. Schmidt-Nielsen, 1959).Encycl. Universalis, art. Physiologie, vol. XIV, p. 616 a.
où n est le nombre d’osmoles par litre. Le tableau donne quelques valeurs caractéristiques de la concentration osmotique.Une différence de pression osmotique de part et d’autre d’une membrane perméable entraîne au moins un déplacement d’eau (transport osmotique), du milieu le moins concentré vers le plus concentré; éventuellement, les sels dissous migrent en sens inverse jusqu’à ce que l’équilibre soit établi entre les deux côtés. Au vu du tableau, il est évident que de tels mouvements doivent exister chez les êtres vivants.2. Perméabilité des membranes biologiquesLes échanges osmorégulateurs sont assurés chez les organismes vivants par des organes perméables spécialisés: tégument, branchies, tube digestif, poumons, reins, etc., constitués par des épithéliums simples ou pluristratifiés, que l’on qualifie indistinctement de membranes biologiques (ce terme englobant en fait, également, les membranes cellulaires). On ne peut entrer ici dans le détail de l’importante question de leur perméabilité, dont les propriétés fondamentales ont été mises en évidence par des travaux effectués sur la cellule végétale (W. Pfeffer, C. E. Overton). Les parois ne sont pas semi-perméables, c’est-à-dire qu’elles se laissent traverser à la fois par l’eau et par les substances dissoutes; mais leur perméabilité est très sélective; ainsi, les ions Na+ et Cl- sont essentiellement extracellulaires, tandis que le potassium (K+) est surtout intracellulaire. L’utilisation, à présent généralisée, des marqueurs radioactifs met en évidence que les échanges transmembranaires sont bidirectionnels, c’est-à-dire qu’ils s’effectuent dans les deux sens. Ils sont, en outre, très intenses, ce qui montre que l’eau et les sels des êtres vivants se renouvellent à grande vitesse.Dans certains cas, ces échanges sont passifs , c’est-à-dire que, pour la substance considérée, leur intensité est prévisible d’après des lois physico-chimiques simples: elle est calculable d’après la différence de concentration (gradient chimique) et la différence de potentiel électrique (gradient électrique) existant entre les deux faces; l’équilibre est régipar la loi de Gibbs-Donnan. Dans d’autres cas, l’équilibre que l’on constate de fait apparaît comme un «défi» permanent à ces mêmes lois; il n’est donc maintenu qu’au prix d’une dépense énergétique que la cellule transporteuse effectue en permanence; on dit que le transport est actif . Ce dernier cas correspond à des « pompes biologiques», dont la plus répandue et la plus étudiée est la «pompe à sodium», qui refoule les ions Na+ hors des cellules [cf. MEMBRANES CELLULAIRES].Les mécanismes biochimiques qui régissent ces transports sont encore mal connus. Dans un certain nombre de cas, on sait qu’ils sont contrôlés par des hormones et par des enzymes (telles que les ATPases, qui libèrent de l’énergie aux dépens de l’ATP), assurés par des transporteurs spécifiques, enfin conditionnés par la structure des membranes (replis de la paroi cellulaire, richesse en mitochondries, etc.).3. Types d’osmorégulationL’équilibre est isosmotique lorsqu’il y a égalité de pression osmotique entre le milieu inférieur et le milieu externe. Pratiquement, ce cas ne s’observe que pour des animaux marins. L’organisme ne tend ni à s’hydrater, ni à se déshydrater. La régulation osmotique porte essentiellement sur les mouvements d’ions (ionorégulation), tandis que le transport osmotique de l’eau est pratiquement nul.Dans les autres cas, l’équilibre est anisosmotique . Quand le milieu intérieur est le moins concentré (hypo-osmotique ), l’animal tend à se déshydrater et à être envahi par le sel qui diffuse à travers les surfaces externes. Il doit compenser ces mouvements en absorbant du liquide externe et en excrétant activement l’excès de sel. Au contraire, si le milieu est le plus concentré (hyperosmotique ), l’animal tend à s’hydrater et à perdre du sel passivement; il compense en excrétant l’excédent d’eau et en absorbant activement des ions à partir du milieu environnant.Les animaux sténohalins sont adaptés à un seul milieu et ne supportent en général que de faibles variations de la salinité externe. En revanche, les euryhalins résistent à de larges variations; les plus caractéristiques sont les migrateurs, tels que l’anguille, le saumon et certains crabes, qui peuvent passer alternativement de l’eau douce à l’eau de mer et chez lesquels les mécanismes osmorégulateurs doivent donc s’inverser quand l’animal change de milieu.L’habitat aérien se caractérise par sa teneur en vapeur d’eau atmosphérique (humidité) et non plus par la salinité. Le problème majeur pour l’animal est de réduire, et d’équilibrer convenablement, les pertes d’eau résultant de l’excrétion, de l’évaporation cutanée et de la vaporisation au niveau des organes respiratoires.Ces divers cas sont illustrés ci-dessous au moyen d’exemples pris dans les différents milieux naturels.Osmorégulation en milieu salinPar leur masse, les océans constituent le milieu d’habitat le plus vaste. La composition et les caractères physico-chimiques y sont remarquablement constants [cf. OCÉANS ET MERS]. La salinité est voisine de 34 g de chlorure de sodium pour 1000, ce qui correspond à une osmolarité de l’ordre de 1 osmole par litre. Assez curieusement, bien que les teneurs absolues en ions minéraux soient très variées dans les liquides extracellulaires des animaux, les proportions de ces ions y sont cependant comparables à celles que l’on trouve dans l’eau de mer. Cette constatation est un argument d’ordre physiologique avancé à la fin du siècle dernier en faveur de l’hypothèse de l’origine marine des êtres vivants (F. Botazzi).Les autres milieux salins naturels sont constitués, d’une part, par les eaux saumâtres, dont la salinité, inférieure à celle de la mer, est très variable; d’autre part, par les salines des déserts où, du fait d’une évaporation intense, la concentration est si élevée que le sel précipite.Osmorégulation des InvertébrésLa grande majorité des Invertébrés sont en équilibre isosmotique avec l’eau de mer, mais la composition de leur milieu intérieur en diffère sensiblement et de façon variable suivant les espèces. Ainsi, chez la ligie (Crustacé isopode), le sodium de l’hémolymphe est de 20 p. 100 plus élevé que dans la mer, tandis que le calcium est multiplié par 3 et le magnésium diminué de 60 p. 100. Le tégument et les autres membranes externes sont très perméables à l’eau et aux sels, comme le montrent les études isotopiques; ce fait semble expliquer qu’ils soient isosmotiques. Les capacités osmorégulatrices peuvent être testées en soumettant ces animaux à des changements plus ou moins amples de la salinité. La figure 1 illustre les types fondamentaux d’osmorégulation observés chez le groupe le mieux étudié, celui des Crustacés. Chez les espèces osmoconformes , l’osmolarité interne suit passivement et égale celle du milieu externe (Maia ); ces animaux se laissent traverser par l’eau et le sel; leur tolérance aux changements de salinité n’est limitée que par la résistance cellulaire; il survient en milieu dilué, mais pas en eau douce; ils se comportent dans une certaine mesure comme des osmomètres. Carcinus est osmoconforme en milieu concentré et osmorégulateur en milieu dilué, ce qui lui permet, à la limite, de survivre en eau douce. Pachygrapsus possède une bonne osmorégulation à toutes les salinités. Le cas le plus impressionnant est celui de l’artémie des salines (Artemia salina ) qui maintient une concentration interne basse, même dans des eaux saturées en chlorure de sodium.Osmorégulation des VertébrésLes Cyclostomes et les Élasmobranches sont en équilibre isosmotique avec l’eau de mer, mais suivant des mécanismes différents. Chez les premiers, qui constituent le groupe le plus primitif, la forte concentration interne est réalisée comme chez les Invertébrés, par l’abondance des ions minéraux, surtout Na+ et Cl-. Chez les seconds, ces ions ne comptent que pour un tiers environ de l’osmolarité plasmatique; les deux autres tiers sont assurés, d’une façon très originale, par de l’urée, qui est un déchet normal et non toxique du métabolisme azoté chez les Élasmobranches; l’urée ne traverse pas l’épithélium branchial et n’est pas excrétée par les reins; de ce fait, elle s’accumule dans le milieu intérieur.Les Poissons téléostéens et les Tétrapodes sont tous hypo-osmotiques à l’eau de mer; leur concentration interne, voisine de 300 milliosmoles par litre, n’est que le tiers de celle du milieu externe. Le cas classique du Téléostéen marin, représenté dans la figure 2, constitue un bon exemple de régulation hypo-osmotique en milieu salin. De tels animaux boivent plus ou moins d’eau de mer, soit pour compenser la déshydratation périphérique, soit accidentellement pour prendre leur nourriture. L’eau de mer est en partie absorbée à travers l’épithélium digestif, et avec elle des ions monovalents (Na+, Cl-, K+); les ions divalents (Ca2+, Mg2+, S42-) sont peu absorbés et sont évacués par l’anus. Ce faisant, un excès de sel est introduit dans l’organisme. Seuls les Mammifères marins ont, comme les autres Mammifères, un rein hautement évolué, pourvu d’anses de Henlé, qui leur permet de fabriquer une urine plus concentrée que le plasma, donc d’éliminer ce sel superflu. Les autres groupes de Vertébrés sont pourvus d’un rein plus primitif, sans anses de Henlé, produisant une urine au mieux isosmotique au plasma. Le relais est alors pris par d’autres organes osmorégulateurs, qui excrètent activement les ions indésirables. Chez les Invertébrés et chez les poissons marins, ces organes sont les branchies. Chez les Tétrapodes autres que les Mammifères, cette fonction est assurée par des glandes nasales ou orbitaires logées dans la tête. La sécrétion de ces glandes, plus concentrée que l’eau de mer, s’écoule soit au niveau des yeux, sous forme de «larmes», soit à l’extrémité du bec. Cela explique notamment que les oiseaux marins soient capables de rester longtemps au large, sans accès à l’eau douce (K. Schmidt-Nielsen).Osmorégulation en eau douceLes eaux douces ont une osmolarité négligeable (quelques milliosmoles) et réalisent un milieu d’adaptation éminemment variable dans sa concentration, sa température, etc. Tous les animaux d’eau douce sont hyperosmotiques au milieu et tendent à gagner du poids et perdre des électrolytes. L’ingestion d’eau par le tube digestif est donc nuisible et, de fait, normalement limitée. En revanche, ils évacuent de l’eau sous forme d’une urine généralement abondante et très diluée par rapport au milieu intérieur. En outre, ils absorbent activement des ions à partir du milieu externe par l’intermédiaire d’organes perméables spécialisés: branchies, papilles anales de larves d’insectes... Un cas typique de cette osmorégulation est celui du téléostéen d’eau douce (fig. 2).Le cas des poissons migrateurs a suscité depuis longtemps beaucoup d’intérêt, notamment parce que leur épithélium branchial peut effectuer suivant le milieu soit une excrétion, soit une absorption des ions, toutes les deux actives. Le déterminisme de cette inversion du transport n’est, à vrai dire, guère élucidé. Ces échanges semblent se produire au niveau de cellules particulières (dont les «cellules à chlorures»), paraissent dépendre d’enzymes membranaires et sont contrôlés par des hormones de plusieurs glandes (hypophyse antérieure, neurohypophyse, hormones stéroïdes, thyroïde).Osmorégulation en milieu aérienEn milieu aérien, la difficulté principale à surmonter réside dans la déperdition d’eau vers l’atmosphère, due à l’évaporation tégumentaire et aux diverses formes d’excrétion. Ces pertes d’eau atteignent leur maximum dans les déserts, où l’air est particulièrement sec. Les surfaces respiratoires, qui sont relativement vastes et, de ce fait, sujettes à l’évaporation, se recouvrent de replis tégumentaires ou s’enfoncent à l’intérieur du corps (Insectes, Vertébrés). Les pertes sont compensées par l’eau de boisson, dont l’ingestion obéit à un déterminisme physiologique rigoureux (réflexe de la soif). Mais l’eau n’est pas toujours disponible à volonté, d’où la nécessité de palliatifs. Certains insectes (Thermobia ) possèdent la propriété remarquable d’absorber activement de l’eau-vapeur atmosphérique, même dans un milieu relativement sec (50 p. 100 d’humidité relative). Cela suppose un travail «contre-osmotique» considérable, dont le mécanisme fait encore problème. D’autres font appel à l’eau métabolique qui se forme par oxydation intracellulaire des substrats énergétiques (par exemple à partir du glucose):C6 H12 6 + 626C2 + 6H2O(cf. MÉTABOLISME, OXYDORÉDUCTIONS BIOLOGIQUES). Chez les Vertébrés, les Amphibiens restent inféodés à l’eau; leur peau nue étant perméable, ils accumulent une quantité importante d’urine très diluée dans leur vessie lorsqu’ils vivent en milieu sec; cette eau de réserve peut être réabsorbée en cas de sécheresse.Les Reptiles et les Oiseaux sont pourvus de phanères épais et diminuent leurs pertes d’eau en excrétant une faible quantité d’urine; cette dernière particularité tient au fait que, chez ces animaux, le métabolisme des matières azotées n’aboutit pas à la formation d’urée (dont l’excrétion entraînerait beaucoup d’eau), mais d’acide urique, qui précipite et donne à l’urine son aspect blanchâtre et sa consistance presque solide.Les Mammifères, enfin, dont le rein est pourvu d’anses de Henlé, peuvent seuls produire une urine au moins deux fois plus concentrée que le plasma; cela leur permet, d’une part, de se débarrasser de leur sel en excès, d’autre part, d’excréter de l’urée comme déchet azoté, donc d’avoir une urine relativement abondante sans inconvénient pour leur équilibre osmotique. Le cas le plus spectaculaire est celui des Mammifères désertiques (le rat-kangourou et le chameau).Le petit rongeur américain Dipodomys , ou rat-kangourou, vit sans eau alimentaire. L’apport d’eau est constitué par l’eau métabolique produite par la combustion des réserves organiques du corps. L’utilisation de 1 g de graisse libère 1,1 ml d’eau. Les pertes périphériques d’eau sont très limitées, d’une part parce que cet animal ne possède pas de glandes sudoripares, d’autre part parce qu’il évite la chaleur du jour et ne cherche sa nourriture que par les nuits fraîches. Son rein pourvu d’anses de Henlé très longues fabrique une urine extrêmement concentrée puisqu’elle atteint 6 osmoles par litre, soit environ = 漣 11 0C. Un tel animal peut boire de l’eau de mer sans aucun inconvénient, ce que l’homme ne peut faire.Le chameau demeure l’exemple le plus populaire. Il peut rester des jours sans boire; à l’inverse, il est capable d’ingurgiter en une seule fois l’équivalent du tiers de son poids en eau. Contrairement à ce qu’affirme la légende, il n’emmagasine de l’eau ni dans sa bosse, ni dans son rumen, ni dans d’autres organes. La bosse est formée de graisses qui, oxydées, libèrent de l’eau métabolique. Mais cette eau n’est pas de grande utilité, car le chameau a une activité diurne. En fait, les performances exceptionnelles du chameau proviennent du fait que cet animal peut tolérer d’une part une forte déshydratation, d’autre part une élévation de plusieurs degrés de sa température interne. Son osmorégulation et sa thermorégulation sont relativement labiles, ce qui explique sa résistance. Ce dernier exemple, emprunté à un organisme pourtant très évolué, montre bien que l’osmorégulation peut fort bien s’accommoder d’assez larges variations des concentrations internes sans que la survie soit compromise.osmorégulation [ɔsmoʀegylɑsjɔ̃] n. f.ÉTYM. Av. 1969; attesté en angl. en 1957; de 2. osmo-, et régulation.❖♦ Physiol. Ensemble des mécanismes par lesquels les organismes maintiennent leur pression osmotique interne dans un équilibre déterminé (qui peut être isosmotique ou anisosmotique) par rapport à celle du milieu externe. || Osmorégulation et ionorégulation. || Osmorégulation intracellulaire. → 2. Osmorégulateur (cit.).0 Chez les Reptiles et les Oiseaux marins, dont le rein est incapable de produire une urine fortement hypertonique et qui cependant ne disposent comme source d'eau et pendant de longues périodes que d'eau de mer, très fortement hypertonique au plasma sanguin, existe une glande originale, apparemment étroitement spécialisée dans les fonctions d'osmorégulation, la glande nasale (K. Schmidt-Nielsen, 1959).Encycl. Universalis, art. Physiologie, vol. XIV, p. 616 a.
Encyclopédie Universelle. 2012.